关键词:新冠肺炎、等温滴定量热、差式扫描量热
摘要
2019 年底在武汉爆发的新冠肺炎 (2019 novel coronavirus disease, COVID- 19) 造成国内外乃至全球的大冲击,无论是医药、经济、贸易及各产业体系均受到巨大考验。藉由全球科学家们日以继夜的努力研究,发现新冠肺炎可能的致病基理;其中较为关键的是新冠病毒感染主要是藉由新冠病毒上的 S 蛋白 (spike protein),专一性的识别攻击人类细胞受体 ACE2 (angiotensin converting enzyme 2) 进而引起后续的反应。相较于同是冠状病毒的 SARS,COVID- 19 识别人类细胞受体的专一性更强高出 20 倍 1 - 3。鉴于新冠肺炎启发,本文将回顾利用等温滴定量热法 (isothermal titration calorimetry, ITC) 及差式扫描量热法 (differential scanning calorimetry, DSC) 研究病毒相关的技术文献。期望能对抗病毒药物开发、疫苗、检验试剂及基础研究等领域,提供有用的信息。
病毒颗粒稳定性及组装
病毒的构造主要以蛋白质外鞘 (衣壳)(capsid),内包核酸性遗传物质 (可能是 RNA 或 DNA,型态则可能是单股或双股)。某些病毒,则在病毒衣壳外围再包有一层脂质套膜 (envelope),且膜上具有一系列的病毒特有的接受蛋白,能专一性识别细胞膜上的受体。因此大部分病毒皆有宿主专一性,能识别宿主细胞表面受体感染特定的目标细胞 (target cell)。例如: 新冠病毒上的 S 蛋白攻击细胞受体 ACE21 - 3、艾滋病毒的 gp120 蛋白专一性识别细胞受体 CD44。
以造成腹泻呕吐症状的诺如病毒为例,他具有高度传染性、传播速度及途径,主要通过摄入受污染的食物或水感染。科学家在研究中制备非传染性病毒颗粒,在结构和免疫学上与完整的病毒相似;该病毒衣壳蛋白主要有两个结构不同的结构域并在体外自组装形成空腔。Ausaret al. 利用 DSC 研究非传染性病毒颗粒在 pH 3 - 8 之间的变化,结果显示病毒颗粒在 pH 3 的条件下能检测到两个分离良好的吸热峰,分别是在 66°C 的热峰及在 92°C 较小的峰;在低 pH 值时,较高温度的热峰会往较低温度转移,而在中性 pH 值时,热峰会叠加在较低温度的热峰 5。
由蛋白水解的研究表明,出现在较低温的热峰是由于蛋白酶稳定 C 端子域,而在较高温的热峰则来自可消化的 N 端子域。这些研究结果与病毒能在酸性条件下保持完整状态的需求一致,病毒在穿透胃细胞膜后会分解并释放其 RNA 并进入(中性 pH)细胞质 6。而由 DSC 研究冠状病毒的蛋白外壳则表现出不同的结构及稳定性以响应环境变化 6。 DSC 结果表明其构象变化分两个步骤进行,将外壳加热到 46°C 时会导致蛋白质的 N 端部分外化,随后升温至 75°C 时其外壳分解成单体蛋白。
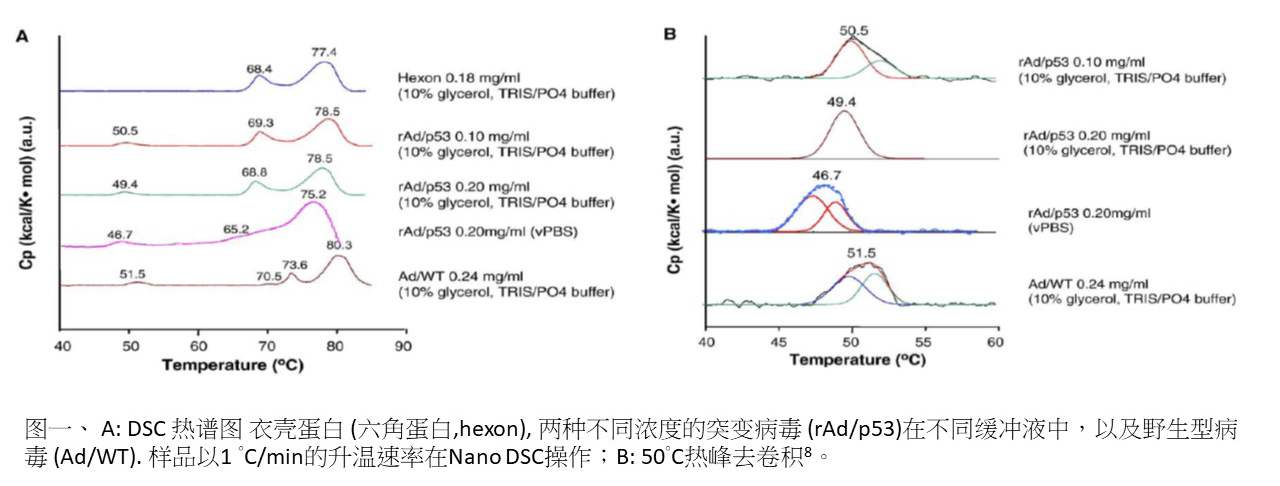
DSC 还被用来调查 pH 值和糖等环境因素对腺病毒热稳定性的影响,而腺病毒是向体细胞提供转基因材料的有利载体 7 - 8。Ihnat et al. 研究了纯化的野生型病毒衣壳蛋白(六角蛋白),包含野生型病毒 (Ad/WT) 和突变病毒 (rAd/p53) 在各种不同缓冲液中 8。结果显示突变病毒表现出三个吸热峰,而野生病毒则具有四个吸热峰。其中衣壳蛋白具有两个吸热峰(如图 1 -A),相较于完整病毒缺少了在 50°C 吸热峰。病毒颗粒中约在 50°C 吸热峰(图 1 -B)的去卷积表明,这种吸热反应是由两个与浓度相关的热效应所组成。结合其他热量数据,作者总结出在 50°C 发生的热效应代表病毒颗粒对蛋白质和核酸的干扰,而大约在 70 及 75°C 的热效应则是来自于六角蛋白的构象变化。并且 50°C 热效应是不可逆的,其位置、大小和形状取决于浓度和环境,因此建议该出峰的位置可用于区分腺病毒突变体,并筛选和优化稳定配方 8。
复杂病毒(如噬菌体及真核病毒)的组装通常需要分子伴侶 (chaperones) 和蛋白酶的帮助。DSC 被长期用來研究病毒的热稳定性,在成熟过程中热稳定性的变化与 X 射线散射和电子显微镜观察到的结构变化相关。也已经在噬菌体 T4、P22、HK97 获得了非常详细的热谱 9 - 11。以 HK97 為例,前体衣殼蛋白 (gp5) 會自组装形成五角和六角蛋白。反过来自组装成前體衣殼頭部 (procapasid),隨後 N 端蛋白水解、衣殼膨胀、构象重组、共价交联和 DNA 的插入等程序。成熟过程能更稳定的病毒顆粒,并且某些步骤(蛋白酶和交联)可防止过程逆转 10。
病毒进入细胞
病毒是通过一种或两种机制的组合进入细胞;一种是与细胞膜直接融合(鞘膜病毒),通过内膜或其他囊泡(非鞘膜病毒)进入。其中后一种机制是由细胞表面受体介导进入。而微热量学已用于研究这两种类型的感染途径。
鞘膜病毒通过与宿主细胞膜融合进入宿主细胞,是由病毒衣殼糖蛋白的构象变化所引發。而构象变化可以由内体 (endosome) 的酸性环境或与细胞表面的特定受体相互作用所触发;例如流感病毒能在酸性条件下与细胞膜融合。病毒外膜是由脂质膜和三个跨膜蛋白组成,其中一种是血凝素,在酸性 pH 值下发生较大的构象变化。这暴露的疏水片段,能驱动病毒与宿主细胞膜融合。在中性酸鹼值的條件下,纯化血凝素的 DSC 熱譜圖觀察到 Tm 值約在 66 °C 的有個較大的吸熱峰,然而在 pH5 时觀察到在 45 °C 具有個大约三分之一熱焓变化的吸熱峰,且蛋白质以非協同去捲積的方式展开 12。結合 DSC 和光谱研究流感病毒,以确定血凝素蛋白嵌入病毒外膜是如何受到 pH 值和温度的响应,結果也显示了在酸性及中性酸鹼值環境下具有顯著的差異 13。与纯化蛋白一样,完整病毒中的血凝素在中性 pH 值條件下在约 66°C 觀察到协同去捲積;而在 pH5 时,Tm 值下降至約 60°C 且變性熱焓基本保持不变,尽管協同作用显著降低。作者认为,酸性環境血凝素稳定性的降低,导致病毒/细胞膜融合由暴露的疏水區介导,但完整病毒中的血凝素在类似条件下比纯化蛋白不稳定度低。在等溫滴定量熱 (ITC) 實驗中,分別研究在中性/酸性環境下对完整及灭活流感病毒進行調研,結果能分辨與細胞膜结合或膜融合,并證實膜融合是仅在酸性 pH 值條件下才會发生的一种吸熱反应 14。
ITC 和其他关于口腔炎病毒的研究表明,在 pH 6.0 时病毒能与磷脂酸胆碱/磷脂酰丝氨酸 (1:3) 的囊泡结合并融合整体呈现放热反应;然而在 pH 7.5 时只有发生结合,但没有发生融合 15 - 16。事实上若是放热反应,说明它可能是由静电作用力驱动的结合反应。当囊泡不含磷脂酰丝氨酸,或者当与病毒融合蛋白结合区域对应的肽中组胺酸被丙胺酸取代时,并未观察到任何结合反应。ITC 数据表明,病毒融合蛋白与细胞结合时,组氨酸残基与带负电的磷脂酰丝氨酸头基非常接近,导致组胺酸质子化并会让融合蛋白与细胞膜结合。
艾滋病毒也许是研究最广泛的包膜病毒。当开始感染时,病毒外壳糖蛋白 (gp120) 与目标细胞上的 CD4 受体相互作用,进而触发病毒包膜的构形变化并导致与靶细胞融合。ITC 对 gp120 和 CD4 相互作用的研究表明 4,是形成大量氢键和范德瓦尔力的焓驱动结合反应 (约- 264 kJ/mol)。但是,较大的结合熵变值很大且不利于反应,表明在结合上存在显着构象自由性下降,导致中等强度的结合 (Ka = 2 x 108 M- 1)。此外,对 CD4 蛋白的独立研究发现其结构在结合时基本保持不变,说明所得到的结合熵主要产生于 gp120 的结构重组。较缓慢的结合速率与复合物形成过程中发生的显着构象变化一致。
对非包膜病毒–猿猴空泡病毒 40(simian virus 40) 的广泛研究,揭示了病毒如何识别和结合宿主细胞上的受体,然而结合得不太紧密导致复制的病毒不能迁移到新的宿主细胞。ITC 数据显示该病毒识别其糖蛋白受体具有毫摩尔级别的亲和力,但聚糖数组筛选实验表明,病毒会优先结合在受体的支链低聚糖。而由晶体学数据分析发现寡糖的两个支链会同时与与结合缝作用,证实了结合的特异性 17。
總結
结合 DSC 和 ITC 研究来了解病毒的结构和作用模式的研究机构正在迅速增长。 DSC 对于阐明病毒结构的热力学轮廓非常宝贵,有助于了解病毒成分的稳定性以及病毒株的可靠性和识别能力。病毒毒性取决于病毒与结合目标的专一性及感染细胞的能力,而 ITC 已被证实是非常强大的检测技术,可用于有效地表征病毒与其受体间发生的任何反应。由于病毒只由几个主要的蛋白质和核酸组成,因此这些成分是如何组装及相互作用,都能藉由微量热技术针对完整的病毒进行热量研究。同时,经由精心设计的 DSC 或 ITC 实验所得到完整病毒特定结构信息,可与其他需要纯化蛋白质或核酸成分才能提供可解释数据的技术相媲美。 快速且准确地评估病毒结构和毒性等因素,对于扩大病毒的结构-活性关系的数据库是至关重要的,从而才能够及时应对病毒爆发和大流行。
参考文献
[1] Meng T, Cao H, Zhang H, Kang Z, Xu D, Gong H, et al. (2020) The insert sequence in SARS-CoV- 2 enhances spike protein cleavage by TMPRSS. bioRxiv. 2020:doi: https://doi.org/ 10.1101 / 2020.02.08.926006.
[2] Hoffmann M, Kleine-Weber H, Krüger N, Müller M, Drosten C, Pöhlmann S. (2020) The novel coronavirus 2019 (2019 -nCoV) uses the SARS-coronavirus receptor ACE2 and the cellular protease TMPRSS2 for entry into target cells. bioRxiv. 2020:doi: https://doi-org-tu.vtrus.net/ 10.1101 / 2020.01.31.929042.
[3] Wan Y, Shang J, Graham R, Baric RS, Li F. (2020) Receptor recognition by novel coronavirus from Wuhan: An analysis based on decade-long structural studies of SARS. J Virol. 2020: doi: 10.1128 /JVI.00127 - 20.
[4] Myszka, D. G., Raymond W. S., Preston Hensley, Michael B.-B., Kwongi, P. D., Hendrickson, W. A., Wyatt R., Sodroski, J., and Doyle, M. L. (2000). Energetics of the HIV gp120 -CD4 binding reaction. PNAS 97, 9026 - 9031.
[5] Ausar, S., T. Foubert, M. Hudson, T. Vedvick and C. R. Middaugh (2006). Conformational stability and disassembly of Norwalk virus-like particles. J. Mol. Biol. 281, 19478 - 19488.
[6] Carreira, A., M. Menendsz, J. Reguera, J. M. Almendral and M. Mateu (2004). In vitro disassemably of aparvovirus capsid and effect on capsid stability of heterologous peptide insertions in surface loops. J. Mol. Biol. 279, 6517 - 6525.
[7] Rexroad, J., C. Wiethoff, A. Green, T. Kierstead, M. Scott and C. R. Middaugh (2002). Structural stability of adenovirus type 5. J. Pharma. Sci., 92, 665 - 678.
[8] Ihnat, P. et al. (2006). Comparative thermal stabilities of recombinant adenoviruses and hexon protein. Biochim. Biophys. Acta 1726, 138 - 151.
[9] Ross, P. D., L. W. Black, M. E. Bisher and A. C. Steven (1985). Assembly-dependent conformational changes in a viral capsid protein. Calorimetric comparison of successive conformational states of the gp23 surface lattice of bacteriophage T4. J. Mol. Biol. 183, 353 - 364.
[10] Ross, P., N. Cheng, J. Conway, B. Firek, R. Hendrix, R. Duda and A. Steven (2005). Crosslinking renders bacteriophage HK97 capsid maturation irreversible and effects an essential stabilization. EMBO J., 24, 1352 - 1363.
[11] Galisteo, M. L. and J. King (1993). Conformational transformations in the protein lattice of bacteriophage P22 procapsids. Biophys. J. 65, 227 - 235.
[12] Remeta, D. P., M. Krumbiegel, C. Minetti, A. Puri, A. Ginsburg and R. Blumenthal (2002). Acid-induced changes in thermal stability and fusion activity of influenza hemagglutinin. Biochemistry 41, 2044 - 2054.
[13] Epand, R. M. and R. F. Epand (2002). Thermal denaturation of influenza virus and its relationship to membrane fusion. Biochem. J. 365, 841 - 848.
[14] Nebel, S., I. Bartoldus and T. Stegmann (1995). Calorimetric detection of influenza virus induced membrane fusion. Biochemistry 34, 5705 - 5711.
[15] Carneiro, F, M. L. Bianconi, G. Weissmuller, F. Stauffer and A. Da Poian (2002). Membrane recognition by vesicular stomatitis virus involves enthalpy-driven protein-lipid interactions. J. Virol. 76, 3756 - 3764.
[16] Carneiro, F. et al. (2006). Probing the interaction between vesicular stomatitis virus and phosphatidylserine. Eur. Biophys. J. 35, 145 - 154.
[17] Neu, U., K. Woeliner, G. Gauglitz and T. Stehle (2008). Structural basis of GM1 ganglioside recognition by simian virus 40. PNAS 105, 5219 - 5224.
Christin T. Choma, 林明申 (Mark Lin), TA Instruments
TA Instruments - Waters LLC
上海市漕河泾开发区钦州北路 1188 号科汇大厦 1601,200233
电话 021 - 34182000
传真 021 - 64951999
info@tainstruments.com
www.tainstruments.com.cn
图片来源:TA 仪器




评论