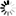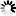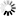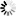

产品详情
文献和实验
相关推荐
提供商 :云舟生物科技(广州)股份有限公司
服务名称 :提供定制载体
此价格为VectorBuilder该类型产品标准服务价格,如需个性化定制,请询价。概述
VectorBuilder的慢病毒诱导基因表达载体将高效的第三代慢病毒载体系统与Tet-On诱导基因表达系统相结合,帮助您实现四环素诱导基因表达盒与宿主基因组的永久整合。
Tet-On诱导基因表达系统是控制哺乳动物细胞中目的基因(GOI)表达时间的有力工具。我们的Tet-On诱导基因表达载体设计用于在没有四环素及其类似物(例如多西环素)的情况下实现GOI的几乎完全沉默,以及响应添加四环素或其类似物之一(例如强力霉素)的强烈快速表达。这是通过多组分系统实现的,该系统在没有四环素的情况下结合了tTS蛋白的活性沉默,以及在四环素存在下rtTA蛋白的强激活。在没有四环素的情况下,来自TetR(Tet阻遏蛋白)和KRAB-AB(Kid-1蛋白的转录抑制结构域)融合的tTS蛋白与TRE启动子结合,导致基因转录的主动抑制。另一方面,rtTA蛋白来源于突变的Tet阻遏因子和VP16(单纯疱疹病毒病毒蛋白16的转录激活结构域)的融合,仅在四环素存在下与TRE启动子结合以激活基因转录。
慢病毒载体系统是将基因永久引入哺乳动物细胞的高效载体。慢病毒载体首先在大肠杆菌中构建为质粒。对于慢病毒诱导基因表达载体,在载体构建过程中,由驱动GOI的四环素诱导元件(TRE)启动子组成的四环素诱导表达盒放置在两个LTR之间。然后将其与几个辅助质粒一起转染到包装细胞中。在包装细胞内,位于两个长末端重复序列(LTR)之间的载体DNA被转录成RNA,辅助质粒表达的病毒蛋白进一步将RNA包装成病毒。然后将活病毒释放到上清液中,上清液可用于直接感染靶细胞或在浓缩后感染。
当病毒被添加到靶细胞中时,RNA货物被穿梭到细胞中,在那里它被逆转录成DNA,并随机整合到宿主基因组中。在载体构建过程中放置在两个LTR之间的诱导表达盒与病毒基因组的其余部分一起永久插入宿主DNA。
虽然我们的慢病毒诱导基因表达载体包括由驱动用户选择的GOI的TRE启动子组成的诱导基因表达盒,但TRE结合调节蛋白rTS和rtTA必须使用单独的辅助载体提供,以便在四环素存在下实现四环素诱导的基因表达,同时在没有四环素的情况下最大限度地减少泄漏表达。对于慢病毒诱导基因表达载体系统,与多合一载体系统相比,双载体系统在四环素存在下可实现更高水平的转基因诱导。多合一载体由两个连续的表达盒组成:由TRE启动子驱动的GOI和由普遍存在或组织特异性启动子驱动的tTS / rtTA基因。对于慢病毒载体,不建议在每个表达盒的LTR之间放置内部聚腺苷酸化信号,因为这会抑制病毒包装。取而代之的是,单个聚腺苷酸化信号被放置在3'LTR中。因此,来自上游TRE启动子的转录通常通过下游启动子和ORF(tTS / rtTA基因)持续到上游ORF末端。这通常导致下游tTS/rtTA表达的部分抑制,从而阻止在四环素存在下有效诱导基因表达。因此,我们建议将靶细胞与携带TRE驱动的GOI的慢病毒和表达tTS / rtTA盒的慢病毒共转导,以实现最佳的诱导效率。
根据设计,慢病毒载体缺乏病毒包装和转导所需的基因(这些基因由病毒包装过程中使用的辅助质粒携带)。因此,由慢病毒载体产生的病毒具有复制不全的重要安全特征(这意味着它们可以转导靶细胞但不能在其中复制)。
高级表达式: TRE启动子可以在诱导状态下驱动GOI的非常高水平表达。
载体DNA的永久整合:传统转染导致DNA几乎完全瞬时递送到宿主细胞中,因为DNA会随着时间的推移而丢失。这个问题在快速分裂的细胞中尤为突出。相比之下,由于病毒载体整合到宿主基因组中,慢病毒转导可以将基因永久递送到宿主细胞中。
高病毒滴度:我们的慢病毒载体可以包装成高滴度病毒。当通过我们的病毒包装服务获得慢病毒时,滴度可以达到>109每毫升转导单位(图/毫升)。在此滴度下,当使用足够量的病毒时,培养的哺乳动物细胞的转导效率可以接近100%。
非常广泛的向性: 我们的包装系统将VSV-G包膜蛋白添加到病毒表面。这种蛋白质具有广泛的嗜性。结果,来自所有常用哺乳动物物种(甚至一些非哺乳动物物种)的细胞都可以被转导。此外,几乎任何哺乳动物细胞类型都可以转导(例如分裂细胞和非分裂细胞、原代细胞和已建立的细胞系、干细胞和分化细胞、贴壁细胞和非贴壁细胞)。神经元通常不受常规转染的影响,可以很容易地被我们的慢病毒载体转导。与我们的系统包装的慢病毒载体比腺病毒载体(对某些细胞类型具有低转导效率)或MMLV逆转录病毒载体(难以转导非分裂细胞)具有更广泛的向性。
基因递送的相对均匀性:通常,病毒转导可以以相对均匀的方式将载体递送到细胞中。相比之下,质粒载体的传统转染可能高度不均匀,一些细胞接收大量拷贝,而其他细胞接收很少拷贝或没有拷贝。
体外和体内有效性: 虽然我们的载体主要用于培养细胞的体外转导,但它也可用于转导活体动物的细胞。
安全:我们载体的安全性由两个功能确保。一种是将病毒包装和转导所需的基因分割成几个辅助质粒;另一种是在载体积分时5' LTR中的启动子活性自失活。因此,在包装和转导过程中基本上不可能出现具有复制能力的病毒。因此,使用我们的载体的健康风险很小。
技术复杂性: 慢病毒载体的使用需要在包装细胞中产生活病毒,然后测量病毒滴度。与传统质粒转染相比,这些程序在技术上要求很高且耗时。
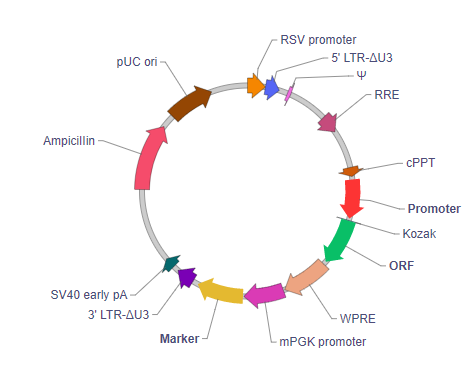
RSV promoter: Rous sarcoma virus promoter. It drives transcription of viral RNA in packaging cells. This RNA is then packaged into live virus.
5' LTR-ΔU3: A deleted version of the HIV-1 5' long terminal repeat. In wildtype lentivirus, 5' LTR and 3' LTR are essentially identical in sequence. They reside on two ends of the viral genome and point in the same direction. Upon viral integration, the 3' LTR sequence is copied onto the 5' LTR. The LTRs carry both promoter and polyadenylation function, such that in wildtype virus, the 5' LTR acts as a promoter to drive the transcription of the viral genome, while the 3' LTR acts as a polyadenylation signal to terminate the upstream transcript. On our vector, 5' LTR-ΔU3 is deleted for a region that is required for the LTR's promoter activity normally facilitated by the viral transcription factor Tat. This does not affect the production of viral RNA during packaging because the promoter function is supplemented by the RSV promoter engineered upstream of 5'LTR-ΔU3 LTR.
Ψ: HIV-1 packaging signal required for the packaging of viral RNA into virus.
RRE: HIV-1 Rev response element. It allows the nuclear export of viral RNA by the viral Rev protein during viral packaging.
cPPT: HIV-1 Central polypurine tract. It creates a "DNA flap" that increases nuclear import of the viral genome during target cell infection. This improves vector integration into the host genome, resulting in higher transduction efficiency.
Promoter: The promoter driving your gene of interest is placed here. Users can select between either the 2nd generation (TRE) or the 3rd generation (TRE3G) Tetracycline-responsive element promoter.
Kozak: Kozak consensus sequence. It is placed in front of the start codon of the ORF of interest to facilitate translation initiation in eukaryotes.
ORF: The open reading frame of your gene of interest is placed here.
WPRE: Woodchuck hepatitis virus posttranscriptional regulatory element. It enhances viral RNA stability in packaging cells, leading to higher titer of packaged virus.
mPGK promoter: Mouse phosphoglycerate kinase 1 gene promoter. It drives the ubiquitous expression the downstream marker gene.
Marker: A drug selection gene (such as neomycin resistance), a visually detectable gene (such as EGFP), or a dual-reporter gene (such as EGFP/Neo). This allows cells transduced with the vector to be selected and/or visualized.
3' LTR-ΔU3: A truncated version of the HIV-1 3' long terminal repeat that deletes the U3 region. This leads to the self-inactivation of the promoter activity of the 5' LTR upon viral vector integration into the host genome (since the 3' LTR is copied onto 5' LTR during viral integration). The polyadenylation signal contained in 3' LTR-ΔU3 serves to terminates all upstream transcripts produced both during viral packaging and after viral integration into the host genome.
SV40 early pA: Simian virus 40 early polyadenylation signal. It further facilitates transcriptional termination after the 3' LTR during viral RNA transcription during packaging. This elevates the level of functional viral RNA in packaging cells, thus improving viral titer.
Ampicillin: Ampicillin resistance gene. It allows the plasmid to be maintained by ampicillin selection in E. coli.
pUC ori: pUC origin of replication. Plasmids carrying this origin exist in high copy numbers in E. coli.









云舟生物科技(广州)股份有限公司
品牌商实名认证
金牌会员
入驻年限:6年