
产品详情
文献和实验
相关推荐
免疫原 :详见说明书
亚型 :IgG
形态 :Lyophilized or Liquid
保存条件 :避光-20℃,保存一年
克隆性 :详询
标记物 :详询(烜雅可供应生物素、酶、荧光素标记的抗体)
适应物种 :h,R,m,r,d
保质期 :一年
抗原来源 :Polypeptide
目录编号 :XY-KT-3017
级别 :生产厂家
库存 :100
供应商 :上海烜雅生物科技有限公司
宿主 :Rabbit,mouse
应用范围 :ih,wb
浓度 :1mg/ml
靶点 :详见说明书
抗体英文名 :TNFβ
抗体名 :TNFβ
规格 :100-200ul
TNFβ抗体产品资料:适应物种:h,R,m,r
级别:生产厂家
纯度:>95%
浓度:1mg/ml
分子量:详见说明书
形态:Lyophilized or Liquid
克隆类型:Polyclonal
规格价格:请直接联系我司客服详询
说明书:请直接向我司客服索取
免疫组化常见问题及对策
免疫组织化学是应用抗原和抗体结合的原理,检测细胞内多肽、蛋白质等大分子物质的 分布。这种方法的特异性强、敏感度高、发展迅速、应用广泛,成为生物学和医学众多学科 的重要研究手段。作为初次使用本公司的抗体从事免疫组化的客户可能遇到许多问题,大体 归纳起来,主要有非特异性着色﹑着色部位不对﹑假阳性﹑无阳性﹑阳性弱五大类以及其他 一些小问题。现在就以上几点分别说明。
非特异性着色
非特异性着色就是在理论着色区域外的着色,这种着色与背景是有区别性的。

主要原因:
- 标本原因,肝肾内源性生物素含量高,与 SABC 结合导致非特异性着色,皮肤肺组织胶 原含量高,由于胶原带负电荷,易吸附试剂导致非特异性着色。
- 切片干涸导致的边缘效应。
- 抗体浓度过高。
- 组织在处理中存在出血区域坏死区。
- 洗涤不充分。
- 显色剂氧化,显色时间长。
- 一抗应做浓度梯度,选择阳性强,背景好,信躁比高的浓度,作为抗体实验浓度;二抗浓 度和时间一般不变。
- 稳定实验条件,严格按操作说明进行。
- 在实验中应保持切片始终处于水平状态避免抗体流失,避免切片干涸。
- 在实验中应将试剂覆盖面积大于切片面积。
- 洗涤要充分,在背景十分深时,可用 37℃预温的 PBS 浸泡
- 防止显色剂的氧化,尤其是配制显色剂使用的容器,最好是灭菌过的;显色时间应在显微 镜下严格控制。
着色部位不对
情况一:本是胞浆抗原有着色,但实验结果显示在胞核有着色。

原因及解决办法:
- 修复时间过长,修复条件十分严苛,这时应降低反应强度,减少修复时间。
- 组织在二中静置时间过长,这时应更换标本。
- 抗体中含有抗核蛋白抗体,这种情况已不多见。
- )标本原因,标本处理不慎会导致总在同一个地方出现着色。
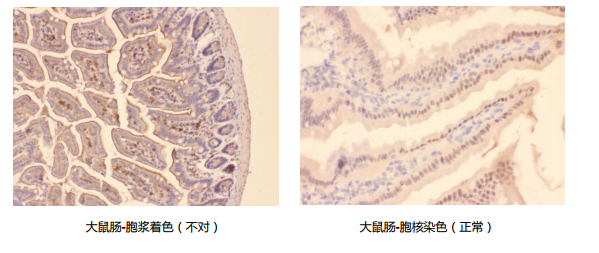
原因及解决办法:
- 核抗原不易暴露,需用热修复或延长修复时间来充分暴露。
- 蛋白质是在胞浆翻译的,再转运至其他部位,所以出现浆着色也属正常。
情况三:本是胞膜抗原但显示结果却是在胞浆和胞膜都有着色。
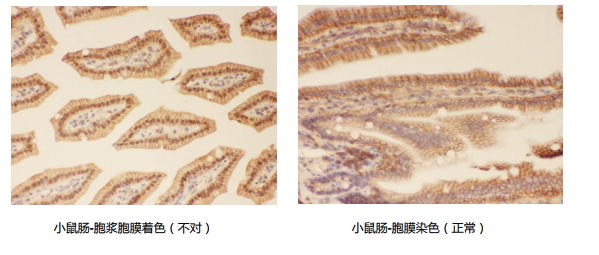
原因:蛋白质是在胞浆翻译的,再转运至其他部位,处于转运过程中的膜蛋白有可能显示在胞浆。
假阳性
如不加一抗也有阳性信号
主要原因:
二抗引起的交叉,球蛋白是一个超家族,哺乳动物种属来缘近的容易发生交叉反应。如:羊 抗兔抗体可与兔标本中球蛋白反应,二抗羊抗小鼠也可与大鼠,小鼠中的球蛋白起反应,尤 其在修复的情况下。
无阳性

主要原因:
1.抗原稳定性问题,由于许多蛋白质半衰期短易被破坏,如 P53 半衰期只有 30 秒而 PCNA 等抗原稳定。
2.标本制作过程中烤片时间过高时间过长,标本制作不规范。
3.实验中漏加试剂,实验操作不规范。
解决方法:
- 在标本固定中应该严格操作,做到及时固定。
- 在浸蜡时温度不要太高,时间不要过长。一般 2 小时×2 次。烤片温度一般在 60℃左右 30 分钟。
- 实验中要严格按照实验步骤操作。在所做指标既有单抗又有多抗时,要将一抗和二抗严格 对应。
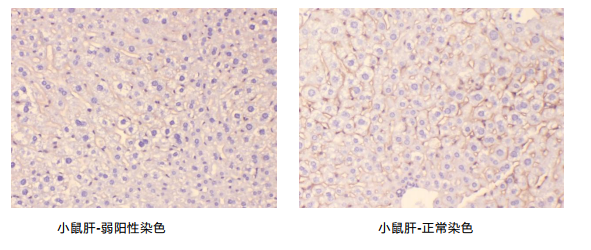
原因及解决方法:
- 一抗浓度过低,解决方法:提高一抗浓度
- 孵育时间过短,解决方法:按说明书严格操作
- 抗体流失,解决方法:补加抗体
- 显色时间过短,解决方法:在显微镜下控制反应时间
- 抗原修复方法不当,抗原修复有许多种方法,如:修复、消化,不消不修,寻找适合的方 法是解决阳性弱的有效方法之一
其他问题及注意事项
- 在实验中尤其在修复后切片容易脱片,切片脱片的原因是什么?
- DAB 中 A 液无颜色或颜色很深是否影响实验结果?
- 冬天气温低洗涤不充分,可能出现背景高, 应对方法用 37℃欲温 PBS 洗涤
- 抗体是否能做兔标本 答:以说明书为准,一般能做人、大鼠、小鼠也可能做兔,这是由于种属亲缘关系决定的
烜雅生物TNFβ抗体热销产品:
| XY-KT-1825 | Anti-PI3K/P85 alpha(Phosphatidyl inositol 3-kinase) 磷脂酰肌醇激酶抗体 |
| XY-KT-1826 | Anti-PIBF(progesterone induced blocking factor) 孕激素诱导阻断因子抗体 |
| XY-KT-1827 | Anti-PIWIL1(piwi-like 1) piwi样1蛋白抗体 |
| XY-KT-1828 | Anti-PKA(Protein Kinase A) 蛋白激酶A抗体 |
| XY-KT-1829 | Anti-PKB/AKT-1(mouse) 蛋白激酶B(小鼠来源抗体) |
| XY-KT-1830 | Anti-PKC(Protein Kinase C) 蛋白激酶C抗体 |
| XY-KT-1831 | Anti-PLAU/uPA (Plasminogen activator,urokinase) 尿激酶型纤溶酶原激活因子抗体 |
| XY-KT-1832 | Anti-PLGF (Placenta growth factor human) 人胎盘生长因子(抗体)相关抗体 YT-0280R(抗大鼠) YT-0234R(抗小鼠) |
| XY-KT-1833 | Anti-PLGF (Placenta growth factor mouse) 小鼠胎盘生长因子抗体 |
| XY-KT-1834 | Anti-PLGF(Placenta growth factor for mouse rat and pig) 胎盘生长因子抗体: 相关抗体:YT-0281R(抗人) YT-0234R(抗小鼠) |
| XY-KT-1835 | Anti-PMP-22(Peripheral Myelin Protein) 外周髓鞘蛋白-22 |
| XY-KT-1836 | Anti-Podoplanin Protein 平足蛋白抗体 |
| XY-KT-1837 | Anti-Pokemon (POK Erythroid Myeloid Ontogenic factor) “扑克蒙”蛋白又称“波克曼”蛋白 |
| XY-KT-1838 | Anti-Pokemon (POK Erythroid Myeloid Ontogenic factor) “扑克蒙”蛋白又称“波克曼”蛋白抗体 |
| XY-KT-1839 | Anti-PP-1 蛋磷酸酯酶-1抗体 |
| XY-KT-1840 | Anti-PP-2A (protein phosphatase 2A) 蛋白质磷酸酶-2A抗体 |
| XY-KT-1841 | Anti-PP-2A(protein phosphatase 2A catalytic subunit) 蛋白质磷酸酶-2A抗体 |
| XY-KT-1842 | Anti-PP2Ac 磷酸酯酶-2Ac抗体 |
| XY-KT-1843 | Anti-PPAR-delta (peroxisome proliferator-activated receptor;PPAR-beta) D型-过氧化酶活化增生受体抗体 |
| XY-KT-1844 | Anti-PPARγ (peroxisome proliferator-activated receptor-gamma) 过氧化酶活化增生受体γ抗体 |
| XY-KT-1845 | Anti-PKB (Akt1/PKB) 蛋白激酶B(兔来源抗体) |
| XY-KT-1846 | anti-P-AKT1/PKB1/2/3(Thr473) 磷酸化蛋白激酶B抗体(丝氨酸磷酸化位点:473) |
| XY-KT-1847 | anti-PKN2 (protein kinase N2) 蛋白激酶N2抗体 |
| XY-KT-1848 | Anti-PR(Progestogerone receptor) 孕激素受体抗体 |
| XY-KT-1849 | Anti-PRL (Prolactin) 鼠抗人泌乳素抗体 |
| XY-KT-1850 | Anti-Prolylhydroxylase 脯氨酸羟化酶抗体 |
| XY-KT-1851 | Anti-PS-1 (Presennillin-1/S182)(NT) 早老素蛋白-1抗体 |
| XY-KT-1852 | Anti-PS-1(Presennillin-1/S182)(CT) 早老素蛋白-1抗体 |
| XY-KT-1853 | Anti-PS-1(Presennillin-1/S182)(NT) 早老素蛋白-1抗体 |
| XY-KT-1854 | Anti-PSA (anti-Prostate Specific Antigen (PSA) , for conjugation and detection) 鼠抗人前列腺特异性抗原抗体 |





上海烜雅生物科技有限公司
实名认证
钻石会员
入驻年限:7年