
产品详情
文献和实验
相关推荐
提供商 :英拜
服务名称 :snoRNA测序
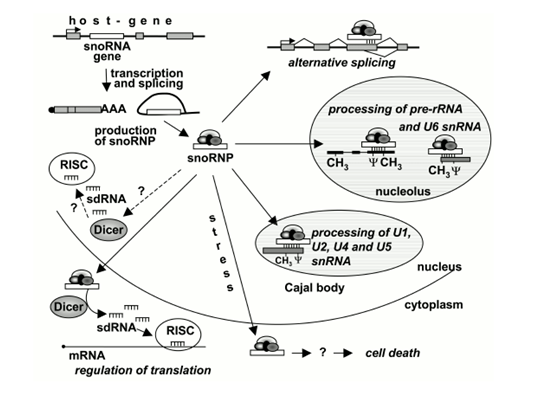
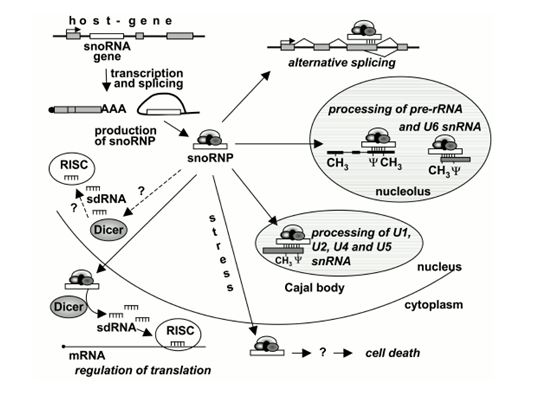
图1 snoRNAs的生物学功能(图片来源:Makarova, J. A., S. M. Ivanova, et al. 2013)
一、实验流程

二、数据分析
1. 测序数据质量控制
2. Mapping统计分析
3. snoRNA表达分析
4. snoRNA差异表达分析
5. 差异snoRNA聚类分析
6. 差异snoRNA热图分析
7. snoRNA的BOX分类
8. snoRNA靶基因分析
9. snoRNA宿主基因预测
三、案例分析
1)“Germline Duplication of SNORA18L5 Increases Risk for HBV-related Hepatocellular Carcinoma by Altering Localization of Ribosomal Proteins and Decreasing Levels of p53” Gastroenterology 2018(IF=20.773)
该研究是基于生殖系拷贝数变异(CNV)的全基因组关联分析(GWAS),通过对比1583例乙型肝炎病毒(HBV)相关的肝癌患者与1540例乙型肝炎病毒(HBV)相关的非肝癌患者,发现了一个低频重复染色体15q13.3与乙型肝炎病毒相关的肝癌患病风险密切相关(P=3.17×10–8;odds ratio, 12.02)。15q13.3的拷贝数与SNORA18L5在肝组织中的表达相关。过表达SNORA18L5的增加了小鼠肝癌细胞的增殖和移植瘤的生长;干扰SNORA18L5降低了肝癌的增殖和肿瘤的生长。SNORA18L5在HepG2和SMMC-7721细胞中的过表达可抑制p53依赖性细胞周期阻滞和凋亡。SNORA18L5的过表达导致核糖体生物合成活性增强,成熟的18S和28S rRNA水平升高,导致核糖体蛋白RPL5和RPL11停留在核仁中,从而阻止它们与MDM2结合。这导致了MDM2介导的p53泛素化和降解的增加。与非肿瘤肝组织相比,HCC组织中SNORA18L5水平升高,且患者生存时间缩短。该研究深入浅出的从中国人口全基因组的生殖拷贝数的突变入手进一步揭示了CNV的突变通过snoRNA的差异表达来影响乙型肝炎病毒(HBV)相关的肝癌。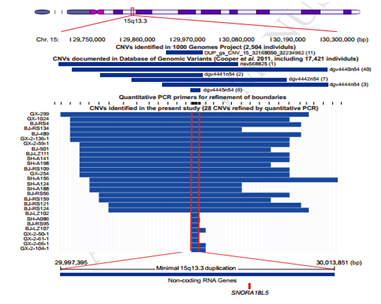
图2 15q13.3的拷贝数突变位点与SNORA18L5相吻合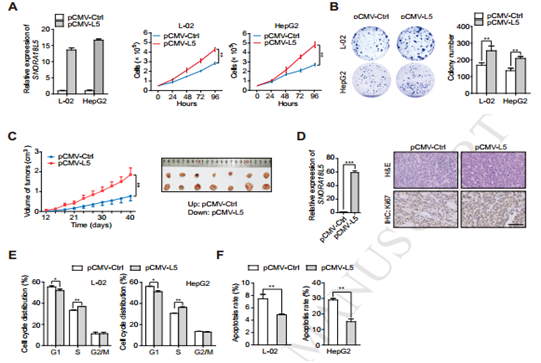
图3细胞与动物实验证明SNORA18L5与肿瘤细胞增殖的关系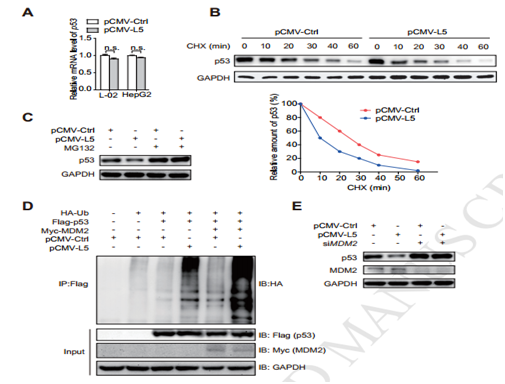
图4 SNORA18L5与MDM2以及抑癌基因P53
2)“AML1-ETO requires enhanced C/D box snoRNA/RNP formation to induce self-renewal and leukaemia”Nature Cell Biology 2017(IF:20.060)
白血病发生需要增强由致癌基因引起的自我更新。其潜在的分子机制尚不完全清楚。本文确定C/D box snoRNAs和rRNA甲基化是白血病干细胞活性的关键决定因素。Aes基因在有融合基因AML1-ETO, PML-RARα和PLZF-RARα的白血病中高表达,但是其功能和机制一直是未知的。研究者通过老鼠模型和细胞系实验,发现Aes对肿瘤细胞的自我更新能力是至关重要的。研究者通过RNA-seq对比Aes(癌基因)缺失前后转录组的变化,意外发现大量的snoRNA下调,而在过表达Aes后snoRNA表达量显著上升,这说明Aes能影响snoRNA的表达。有趣的是,研究者利用CRISPR技术敲除snoRNA后,发现即使是敲除单个snoRNA,rRNA的甲基化显著降低,并且抑制白血病细胞的克隆形成能力。这说明,snoRNA不仅是一类受癌细胞调控的非编码RNA,更参与了癌症的发生发展。AES通过与RNA解旋酶DDX21相互作用诱导snoRNA/RNP形成。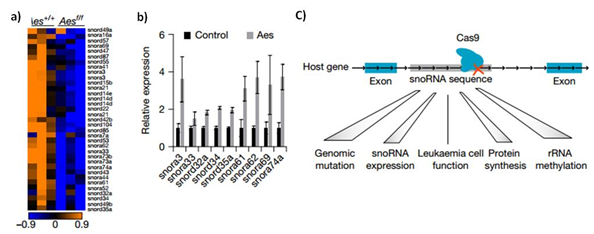
图5 Aes的缺失导致snoRNA的下调
3)“The noncoding RNAs SNORD50A and SNORD50B bind K-Ras and are recurrently deleted in human cancer” Nature genetics 2016 (IF=27.125)
研究人员发现一对原以为只是充当细胞管家的RNA分子,在超过四分之一的常见人类癌症中缺失。研究人员比较了21种不同癌症类型中的5,473个肿瘤基因组及周围正常组织的基因组。研究人员发现在12种常见人类癌症,包括皮肤癌、乳腺癌、卵巢癌、肝癌和肺癌中,10-40%的肿瘤缺失一对称作为SNORD50A/B的snoRNAs。在人类黑色素瘤和肺癌细胞中删除SNORD50A/B时,细胞更快速地分裂,显示出更多的癌性状。
通过人类蛋白微阵列检测到SNORD50A/B能直接结合到KRAS蛋白,当SNORD50A/B结合KRAS时,它会抑制KRAS蛋白结合一种称作为法尼基转移酶(farnesyltransferase)的活化分子的能力。法尼基转移酶改变KRAS蛋白使得它能够去到细胞膜等待外部生长及分裂信号。SNORD50A与SNORD50B缺失能增加GTP结合的数目,激活K-Ras,以及增加法尼基转移酶结合到K-Ras,这些均揭示了K-Ras的突变与SNORD50A与SNORD50B缺失起协同作用。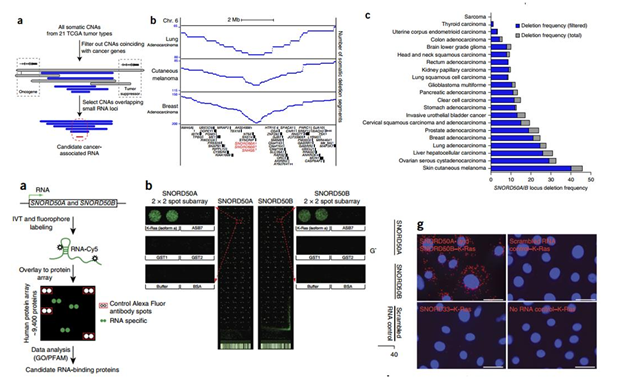
图6 snoRNA多数据库分析以及SNORD50A/B与KRAS蛋白的直接结合
四、研究思路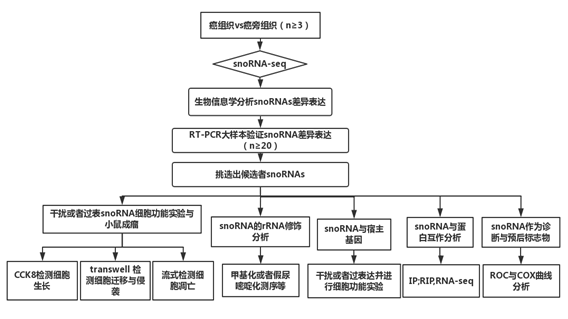
五、研究方向
1.snoRNAs与肿瘤
差异表达的snoRNAs与肿瘤发生发展的功能机制研究;snoRNAs作为肿瘤诊断与预后的分子标志物的分析。
2.snoRNAs与疾病
检测疾病模型组织与正常组织差异性差异表达变化的snoRNAs,结合机制与功能分析,阐明snoRNAs在疾病中的作用机理。
六、送样要求
1. 样品类型:细胞、新鲜组织或RNA样品。
2. 样品量:细胞样品请提供至少5×106个细胞,组织样品请提供至少100mg的组织块或切片,RNA样品请提供10μg以上的总RNA。
3. 样品质量:RNA无明显降解,提取的总RNA OD260/280值在1.8~2.2之间,浓度 ≥ 500ng/μl,28S:18S ≥ 1.5,RIN ≥ 8。
4. 样品保存:细胞样品或新鲜组织块(切成~50mg的小块)可用TRIZOL或RNA保护剂处理或液氮冻存后,-80℃保存。RNA样品可溶于乙醇或RNA-free的超纯水中,-80℃保存。样品保存期间避免反复冻融。
5. 样品运输:样品置于1.5 ml管中,封口膜封好,干冰运输。









上海英拜生物科技有限公司
实名认证
钻石会员
入驻年限:13年